One of the fundamental problems trees have to solve is how to transport water up to the leaves. The ability to solve this problem poses a constraint on the maximum height of trees. The standard freshman physics textbook answer to the question, “What’s the mechanism that lifts the water?”, is capillary action (see Halliday and Resnick for example). Capillary action is the term for the ability of a narrow tube (a capillary) to draw liquid into itself via the adhesive force between the liquid and the tube walls. In plants, the relevant tubes are known as Xylem and the fluid is water. In trees, capillary action alone is not the only mechanism to transport water. First, without evaporation, capillary action would only transport water until mechanical equilibrium is reached and the flow stops. There’s also osmotic pressure at the roots drawing the water in from the soil. There are probably also osmotic effects throughout the tree itself.
Anyway, ignoring all that, let’s see where the freshman physics treatment gets us–what’s the maximum possible height of a tree if capillary action is the limiting factor? To solve this problem, we’d need to think about branching and the fact that pores in the leaves constitute the open ends of the “capillaries”. In the absense of any knowledge about trees, this is a hard problem. Let’s then ignore all the complications and just start with a single capillary tube. We’ll put branching back in to some extent a bit later.
So, what’s the height of a column of water in a single tube with a circular cross-section with radius 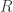


where 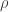

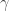
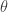

Since 

where the 0.14 cm comes from plugging in lookupable values. The question now remaining is, “what’s 

Okay, so a tree is not composed of a set of parallel tubes of constant diameter from top to bottom. If we bring back in some of the reality of trees, does capillary action postdict something observably true?
Branching should change the story because the height as calculated above is determined by the surface-to-volume ratio of the column of water. If that ratio goes up, the height goes up too. How can we crank the ratio up? If the xylem in upper branches are narrower than those lower down and there are enough of them (is this true? probably), that’d do the trick.
Let’s now model the tree as a very simplified system–a “tree” of height 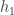



where 

We can now calculate the forces for this model and see what the results might tell us about trees. The upward force—the total due to the adhesive force in each of the N upper tubes—is:

We see that the upward force scales like the square-root of the number of tubes in the upper branches and is independent of the height. The gravitational force is due to the mass of all the water in the branched tree and is given by:

We see that the downward force is sensitive to the overall height of the tree and the location of branching, but it is not sensitive to the number of upper tubes.
Actually making a numerical prediction about the maximum height would be futile here as trees don’t branch only once and I’d have to assume a few more numbers I don’t know, but we can use this analysis to make a qualitative prediction about branching.
Combining the results of Equations (2) and (3) makes a prediction for the structure of a tree if the tree’s structure tries to both maximize the height (as may be the case in dense forrests, where maximizing the height may also maximize the amount of light getting to the leaves) and maximize the amount of water transported simultaneously. The upward force on the water grows like 
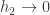
*Here’s how to calculate the packing fraction for circles in a 2D hexagonal close packed (HCP) arrangement. In english, the HCP structure is the “honeycomb nesting”, in which a row of touching circles sits in the interstices of the adjacent rows (consider just the “A” sheet in this picture). The packing fraction,
. The total area covered is the area of the rectangle covered by the M rows of N disks, which is the length of the rows times M times the height per row. The height per row is the same as the distance between the centers of the rows. The centers of 3 adjacent disks mark the vertices of an equilateral triangle with side length 2r. The height of that triangle and thus the distance between rows is
. So, the area of the covered rectangle is
. The packing fraction is the ratio of those areas,
.
There’s probably lots of interesting things about packing of objects. I remember hearing about M&M packing in my undergrad condensed matter course, and that’s pretty fun. The big thing there is oblate spheroids randomly pack a lot tighter than spheres do.
**Do I really believe this is the mechanism behind the branch distribution of the sequoia? I don’t know. It’d help if I actually knew something about trees. It’s at least consistent with this paper that claims that tree branch ratios are all about getting light to the leaves efficiently, where efficiently is defined in the paper..

{kind=link}
{kind=link}